2012年12月30日掲載 【昆虫の変身を抑えるメカニズムの解明 ~優しい殺虫剤の開発に向けて~】
殺虫剤による害虫防除は、安定的な農作物の生産を可能にした一方、その使用を誤ると、環境汚染や人体への悪影響を引き起こす危険性があります。また、ミツバチのような受粉昆虫や害虫の天敵であるテントウムシなどの益虫も殺してしまう場合もあります。そのため、環境や人体にやさしく、受粉昆虫や天敵などの益虫をなるべく殺さない、害虫にだけ選択的に作用する殺虫剤の開発が求められています。私達は、このような条件を満たす殺虫剤を合理的に開発するために、脱皮・変態に関わる昆虫固有のホルモンである幼若ホルモンに着目しています。未だ明らかにされていない幼若ホルモンの分子機構を解明し、新たな殺虫剤の開発に貢献したいと考えています。
昆虫の脱皮と変態
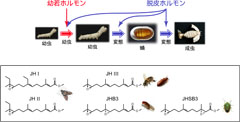
図1: 昆虫の脱皮と変態
幼若ホルモンと脱皮ホルモンが両方作用すると、幼虫から幼虫へと脱皮し、幼若ホルモンがなくなって脱皮ホルモンだけ作用すると、幼虫から蛹、蛹から成虫へと変態します(図上)。
昆虫の種類によって構造が異なる幼若ホルモン(図下)。
カイコガの写真は比留間 潔氏(弘前大学農学生命科学部)より、他の昆虫写真は「昆虫研究所」より借用しました。
(クリックで拡大します)
昆虫は卵からかえると、幼虫から幼虫へとほとんど姿形を変えずに脱皮を繰り返して大きくなります。十分に大きくなると、今度は幼虫から蛹、蛹から成虫へと劇的な「変身(変態)」を行います(図1)。この昆虫の脱皮と変態は、幼若ホルモンと脱皮ホルモンという2つのホルモンによって制御されています(図1)。幼若ホルモンと脱皮ホルモンが両方作用すると、幼虫から幼虫へと脱皮し、幼若ホルモンがなくなって脱皮ホルモンだけ作用すると、幼虫から蛹、蛹から成虫へと変態します(図1)。つまり、幼若ホルモンには幼虫が蛹に変態するのを抑える働き(変態抑制作用)があります。幼若ホルモンは昆虫の種類によって構造のバリエーションが存在するため(図1)、特定の害虫に選択的に作用する殺虫剤を開発する上で、非常によい標的になると考えられます。
幼若ホルモンと脱皮ホルモンの分子機構
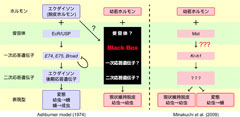
図2: 幼若ホルモンと脱皮ホルモンの分子機構
脱皮ホルモンの分子機構はよく分かっていますが、幼若ホルモンの分子機構はよく分かっていませんでした(図左中)。2009年に我々の研究グループは、その一端を解明し(図中一番右)、今回はMetとKr-h1の関係を明らかにしました(図中の赤???の部分)。
(クリックで拡大します)
脱皮ホルモンが細胞のどの分子に受け取られ(受容体)、どのような遺伝子を誘導・抑制しているのかは、詳しくわかっています(図2)。一方、幼若ホルモンに関しては、その存在が発見されて70年以上経過しましたが、ごく最近まで、受容体のみならず、幼若ホルモンに応答する遺伝子群はほとんどわかっておらず、ほとんどブラックボックスの状態でした(図2)。これまでに私達は、甲虫(カブトムシやテントウムシなど)のモデル昆虫であるコクヌストモドキ(穀物の害虫)を用いて、Krüppel homolog 1(Kr-h1)という遺伝子が、幼若ホルモンによって誘導され、昆虫の変態を抑える働きを持つことを明らかにしました(図2)。また、Kr-h1遺伝子が幼若ホルモンの受容体と考えられているMethoprene tolerant(Met)というタンパク質の下流で機能することも明らかにしました。しかし、幼若ホルモンを受け取ったMetが、Kr-h1遺伝子をどのように働かせているのか、詳しいメカニズムは明らかにされていませんでした。
変態抑制遺伝子Kr-h1の誘導メカニズム
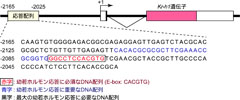
図3: Kr-h1遺伝子の幼若ホルモン応答配列
Kr-h1遺伝子の応答配列(141bp)は、発現する場所(+1)から約2kb上流に位置し、bHLH型転写因子が結合するE-box配列(CACGTG)を有します。
(クリックで拡大します)
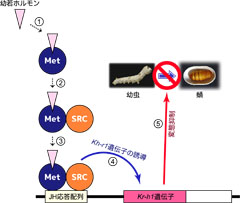
図4: Met/SRCによるJH依存的なKr-h1遺伝子の誘導メカニズム
細胞内に取り込まれた幼若ホルモンは、受容体であるMetに受け取られます。幼若ホルモンを受け取ったMetは、SRCと複合体を形成して幼若ホルモン応答配列に結合し、Kr-h1遺伝子を誘導します。そして誘導されたKr-h1が、幼虫が蛹になるのを抑えます。
(クリックで拡大します)
最近、私達はKr-h1遺伝子が、昆虫の体だけではなく、昆虫の培養細胞でも幼若ホルモンによって誘導されることを発見しました。 さらに、培養細胞を用いた生物発光レポーターアッセイ法(ホタルの光を利用した実験方法)によって、Kr-h1遺伝子の上流域から幼若ホルモンに反応するために必要なDNA塩基配列(JH応答配列)を特定することに成功しました(図3)。特定したJH応答配列は、E-boxと呼ばれるbHLH型転写因子(遺伝子の発現や抑制を調節するタンパク質)が結合するDNA配列を含んでいることが明らかになりました(図3)。
特定したJH応答配列と相互作用するタンパク質(転写因子)を探索したところ、幼若ホルモン受容体であるbHLH-PAS型転写因子Methoprene tolerant(Met)とSteroid receptor coactivater(SRC)を見出すことに成功しました。
これらの結果をもとに、幼若ホルモンによるKr-h1の誘導メカニズムを提唱しました(図4)。
- 1) 細胞に運ばれた幼若ホルモンはMetによって受け取られます。
- 2) 幼若ホルモンを受け取ったMetはSRCと結合します。
- 3) 幼若ホルモン/Met/SRCの複合体は、幼若ホルモン応答配列に結合します。
- 4) 幼若ホルモン/Met/SRC/JH応答配列の複合体が、Kr-h1を誘導します。
- 5) 誘導されたKr-h1が昆虫の変態を抑えます。
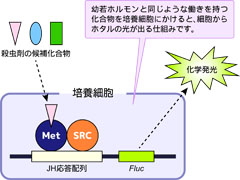
図5: 環境に優しい新規殺虫剤評価システム
培養細胞にMet遺伝子、SRC遺伝子、幼若ホルモン応答配列を導入し、有力な殺虫剤の候補分子を簡単に評価できるシステムを開発しました。
(クリックで拡大します)
興味深いことに、ゲノム情報がわかっている昆虫からMet遺伝子、SRC遺伝子、Kr-h1遺伝子上流のJH応答配列を探索したところ、すべての昆虫にこれらの遺伝子や配列が保存されていました。このことから、Met/SRCによる幼若ホルモン依存的なKr-h1遺伝子の誘導は、昆虫に共通なメカニズムであることが示唆されました。
おわりに
現在私達は、Met遺伝子、SRC遺伝子、JH応答配列を利用して、環境に優しい農薬の開発を目指してさらに研究を進めています。具体的には、害虫のMet遺伝子、SRC遺伝子、幼若ホルモン応答配列を単離し、培養細胞で殺虫剤の候補となる薬剤を簡単に評価できるようなシステムの開発を行っています(図5)。このシステムを農薬メーカーや大学等の研究機関に利用して頂き、環境に優しい殺虫剤を開発していきたいと考えています。
参考文献・サイト
- 該当記事のプレスリリース
- Kayukawa T, Minakuchi C, Namiki T, Togawa T, Yoshiyama M, Kamimura M, Mita K, Imanishi S, Kiuchi M, Ishikawa Y, Shinoda T (2012) Transcriptional regulation of juvenile hormone-mediated induction of Kruppel homolog 1, a repressor of insect metamorphosis. Proc. Natl. Acad. Sci. U.S.A. 109(29): 11729-11734
- Minakuchi C, Namiki T, Shinoda T (2009) Kruppel homolog 1, an early juvenile hormone-response gene downstream of Methoprene-tolerant, mediates its anti-metamorphic action in the red flour beetle Tribolium castaneum. Developmental Biology 325(2): 341-350
« ショウジョウバエは栄養のことを考えて食べる | トピック一覧 | ゴールで生活するアブラムシの快適な住まいづくり »
応用動物学/昆虫学最新トピック
プロの研究者でもまだ知らないような、出来たてホヤホヤの最新研究成果を分かりやすくお伝えします。
日本応用動物昆虫学会(応動昆)
「むしむしコラム・おーどーこん」は、日本応用動物昆虫学会電子広報委員会が管理・運営しています。