2013年12月19日掲載 【GFPよりも使いやすい遺伝子組換えマーカーを目指して】
ショウジョウバエでは遺伝子組換え体を色や形などの見た目で判別できますが、他の昆虫では組換え体の判別はGFPなどの蛍光マーカーで行われています。しかし、蛍光マーカーを用いたスクリーニングは、蛍光顕微鏡が高価であることに加えて、実際やってみると大変骨の折れる作業であることをご存知の方もいるでしょう。
ショウジョウバエ以外の昆虫で遺伝子組換え実験をしている人は、ハエは肉眼で簡単にスクリーニングできるのでうらやましいと一度は思ったことがあるのではないでしょうか。
今回私たちは、様々な昆虫に適用可能で、組換え体を見た目(色)で判別できるマーカーを作り出すことに成功しました。(Osanai-Futahashi et al. 2012)
組換え体のスクリーニングは大変
私が所属する農業生物資源研究所・遺伝子組換えカイコセンター(現遺伝子組換えカイコ研究開発ユニット)は、2001年に田村俊樹博士が開発した遺伝子組換えカイコの作出技術を基盤としています。遺伝子組換え系統は毎年一回飼育して保存しているものだけで数百系統あり、その数は年々増加しています。
私はこの研究室に移ってきて初めて遺伝子組換え昆虫を扱ったのですが、最初に驚いたのが遺伝子組換えカイコのスクリーニングに多大な労力がかかっていることです。カイコはバランサー染色体がないので、毎回組換え体のスクリーニングが必要です。また、ショウジョウバエと比べ飼育に手間がかかるので、早い段階で組換え体を判別する必要があります。そこで、3xP3EGFPなど、眼で蛍光タンパク質を発現するマーカーが使われていましたが、蛍光タンパク質が光りはじめてから2日後には頭が黒く着色して蛍光が見えなくなるため、卵の発生時期に厳密に合わせてスクリーニングを行う必要がありました。さらに、蛍光が意外に見にくいので、慣れていない人が行うと全く光っていない卵が選ばれたりするため、実際のスクリーニングは、経験者か博士号を取得した研究員が行っていました。このため、毎年春の一斉飼育では、全ての研究者が実験をストップさせて大量の保存系統のスクリーニングを行っていました。
私は研究室に入った当初のスクリーニングの印象が大変強く、簡単に組換え体を判別できる方法はないものかと考えました。
メラニン合成系遺伝子の強制発現によるカイコの体色変化
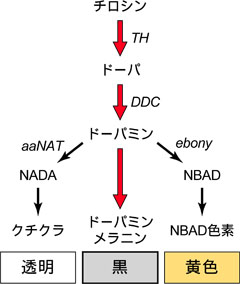
図1: 昆虫のメラニン合成経路。TH, Tyrosine Hydroxylase; DDC, Dopadecarboxylase; aaNAT, arylalkylamine N-acetyltransferase; NADA, N-acetyl-dopamine; NBAD, N-beta-alanyl-dopamine
(クリックで拡大します)
遺伝子組換え体を選別するためには、染色体の片方に遺伝子が導入されただけで判別できる(優性に働く)点が重要になります。カイコの卵の色の変異体に、原因遺伝子を組み込んで判別する手法は存在しましたが(関連記事カイコの変異体から分かったショウジョウバエの眼が黒くない理由を参照)、このような変異体をレスキューする方法は、数多く存在するカイコの系統にそのまま適用することができない点が問題になっていました。このような経緯から、発生の早い段階で働く優性マーカーの開発が切望されていました。
カイコの幼虫をはじめ、ほとんどの昆虫の体表の黒い色は主に「メラニン系色素」から成っており、ドーパミンという物質から合成される「ドーパミンメラニン」が主要なメラニン系色素だと考えられています(図1)。したがって、ドーパミンメラニンの産生を人為的にコントロールできれば、幅広い昆虫に適用可能な優性形質を持つマーカーとして利用できるのではないかと考えました。

図2: Bm-ebony遺伝子、Bm-aaNAT遺伝子を強制発現したカイコ幼虫
(a)上: GAL4/UASシステムにより、Bm-ebony遺伝子を全身で強制発現させたカイコの3齢幼虫。下: ネガティブコントロール。両方とも元のカイコの系統は黒縞である。
(b)上: GAL4/UASシステムにより、Bm-aaNAT遺伝子を全身で強制発現させたカイコの5齢幼虫。中・下: ネガティブコントロール。3頭とも元のカイコの系統は黒縞である。
(c) ▲はBm-aaNAT遺伝子をバキュロウイルス由来のImmediate Early 1(IE1)プロモーターに直接つないだカイコの1齢幼虫
(クリックで拡大します)
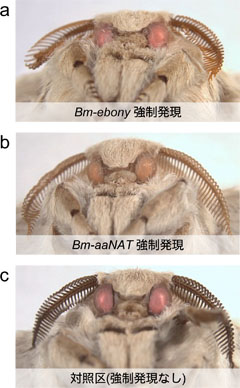
図3: Bm-ebony遺伝子、Bm-aaNAT遺伝子を強制発現したカイコ成虫
(a) GAL4/UASシステムによりBm-ebony遺伝子を強制発現したカイコ成虫
(b) GAL4/UASシステムによりBm-aaNAT遺伝子を強制発現したカイコ成虫
(c) 対照区。(a)と(bのカイコの触角の色が明るい触角に変化している。
(クリックで拡大します)
ドーパミンは、黒いドーパミンメラニン以外にも、透明なクチクラ層(上皮の外側の膜)の合成に必要な物質であるN-acetyl-dopamine (NADA)の合成や、黄色の色素の生成に必要なN-beta-alanyl-dopamine (NBAD)の合成にも使われます(図1)。そこで、NADA、もしくはNBADの合成を強化することにより、ドーパミンメラニンの合成を阻害できるのではないかと考えました。つまり、新たに色をつけるのではなく、既に存在する色をなくしてしまうという方法で組換えマーカーを作るという発想です。まずは、カイコの幼虫でNADA、NBADの合成に関わる酵素を産生する遺伝子を強く働かせることによって、幼虫の体色が変化するかを調べました。
GAL4/UASシステムを用いて、ドーパミンからNBADを合成する酵素の遺伝子(Bm-ebony)を全身で強く発現させてみたところ、予想とは異なり、1齢幼虫の色はほとんど変化しませんでした。しかし、2齢以降では予想通り黒い色が黄色っぽくなるという変化が見られました。色の変化を明確に観察するため、幼虫に黒い縞模様が出る黒縞系統を用いた場合、3齢幼虫では黒い部分のほとんどが黄色になり(図2a)、成虫触角の色も明るい褐色に変化しました(図3)。NBADの合成にはドーパミンのほかに、ベータアラニンが必要であるため、Bm-ebonyの作用の齢による違いは、ベータアラニンの有無が関係しているのかもしれませんが、1齢で判別できないとなると組換えマーカーとして利用するにはもう一歩でした。
そこで次に、NADAを合成する酵素の遺伝子(Bm-aaNAT)をカイコ幼虫の全身で発現させてみました。すると、Bm-aaNATを強制発現させたすべての個体が、1齢幼虫のうちから、体色が黒色から明るい褐色へと変化しました。体色変化は、全ての齢の幼虫の黒い斑紋や成虫の触角でも確認されました(図2b、3)。NADAの合成にはドーパミン以外にアセチルcoAが必要ですが、アセチルcoAは豊富に存在する基質であるため、Bm-aaNATは1齢幼虫でも体色を薄くすることができたことが考えられます。
実際にマーカーとして使用するためにはGAL4/UASシステムでなく、遺伝子を直接プロモーターにつないだときの変化を見る必要があります。そこで、早い段階から遺伝子発現を誘導するバキュロウイルス由来のImmediate Early 1(IE1)プロモーターに直接Bm-aaNAT遺伝子をつなぎ、組換え体を作製してみると、GAL4/UASシステムと同様にすべての個体で体色変化が確認できました(図2c)。心配していた行動異常は見られず、生存率、成長スピードや産卵数もほとんど通常の個体と変わりませんでした。このことから、Bm-aaNAT遺伝子が、遺伝子組換え個体を判別するマーカーとして実際に利用可能であることが分かりました。
Bm-aaNAT強制発現はキイロショウジョウバエとテントウムシの色も変える

図4: 他の昆虫におけるBm-aaNAT遺伝子強制発現の影響
(a) GAL4/UASシステムを用いてBm-aaNAT遺伝子を強制発現したキイロショウジョウバエの成虫(右)は、対照区の個体(左)と比べ、体色が薄く、体毛も白い。GAL4はhsp-GAL4を用いている。
(b)ショウジョウバエhspプロモーターにBm-aaNAT遺伝子を直接つないだコンストラクトを形質転換したナミテントウの幼虫(中、右)。形質転換していない個体(左)と比べ、体色が劇的に薄くなっている。ヒートショック処理を行うと、右個体のようにさらに色が薄くなる。
(クリックで拡大します)
メラニンはほぼ全ての昆虫に存在すると考えられることから、Bm-aaNATの強制発現によりカイコ以外の昆虫の色も変わることが考えられました。そこで、名古屋大学の新美輝幸先生にお願いして、キイロショウジョウバエとナミテントウでBm-aaNAT遺伝子の強制発現を試みました。キイロショウジョウバエはGAL4/UASシステムを使うと成虫で体や毛の色が薄くなりました(図4a)。また、ナミテントウではBm-aaNAT遺伝子をキイロショウジョウバエのheat shock protein 70 (hsp)プロモーターにつないで組換え体を作製していただいたところ、幼虫の黒い色が薄くなる系統が得られました(図4b)。この系統にヒートショックをかけると、ほぼ完全に黒い色素ができなくなったことから(図4b)、hspプロモーターが駆動するBm-aaNAT遺伝子の働きにより体色変化が起きたことが示唆されました。このことから、鱗翅目以外の昆虫でも適切なプロモーターと組み合わせることができれば、Bm-aaNAT遺伝子を遺伝子組換えマーカーとして応用可能であることが分かりました。
以前、海外の研究者と話をしていた際に、THやDDCなどのメラニン合成酵素遺伝子の強制発現で色をつけてマーカーにすることはできないかと聞かれたことがあります。しかし、メラニンなどの色素の合成は複数の酵素の協調的な発現が必要であることが知られており(Futahashi et al. 2010)、1つの遺伝子のみを発現させるだけでは不十分な可能性が考えられます。今回はすでに体表に存在しているドーパミンメラニンを一つの酵素で別の物質に変えるという逆転の発想が功を奏した例といえそうです。
参考文献・サイト
- 該当記事のプレスリリース(農業生物資源研究所)
- 該当記事の日本語要約(NPGネイチャー アジア・パシフィック)
引用文献
- Osanai-Futahashi M, Ohde T, Hirata J, Uchino K, Futahashi R, Tamura T, Niimi T, Sezutsu H. (2012) A visible dominant marker for insect transgenesis. Nat Commun. 3:1295. doi: 10.1038/ncomms2312.
- Futahashi R, Banno Y, Fujiwara H. (2010) Caterpillar color patterns are determined by a two-phase melanin gene prepatterning process: new evidence from tan and laccase2. Evol Dev. 12(2): 157-167.
« 冬の寒さが冬尺蛾の種分化を引き起こす | トピック一覧 | 「生物を生きたまま電子顕微鏡で高解像度観察する」~昆虫が分泌する物質を規範とした"防護服"ナノスーツの開発~ »
応用動物学/昆虫学最新トピック
プロの研究者でもまだ知らないような、出来たてホヤホヤの最新研究成果を分かりやすくお伝えします。
日本応用動物昆虫学会(応動昆)
「むしむしコラム・おーどーこん」は、日本応用動物昆虫学会電子広報委員会が管理・運営しています。