2013年10月25日掲載 【ヒサカキの花蕾を餌とするソトシロオビナミシャクの適応進化】
昆虫の幼虫には翅がなく、ほとんどの場合、成虫のようには移動できません。そのため、幼虫の中には、自分で餌を選べず、産卵された場所が餌となるものもいます。そのような昆虫では、産卵行動が非常に重要なカギを握っています。今回は、花を食べるシャクガの幼虫の餌利用と産卵行動の進化について調べた研究を紹介します。
はじめに
地球上にみられる多くの昆虫は陸上植物とともに、多様化してきたと考えられています。植食性昆虫は、よりよく植物を利用できるように、一方、植物は昆虫に食べられないように様々な進化を遂げています。多くの植物は桜のように雄と雌の機能を併せ持つ両性花をつけますが、少数派ながらも、雄個体と雌個体が分かれている植物もあります。このように個体により性が異なる植物では、成長速度や根の張り方、花の大きさなど様々な形質で性差がみられることが分かってきています。また、葉に残された食害痕の比較からも、食害を受ける量が植物の性により異なる場合があることも報告されています。これらのことから、昆虫にとって、性が異なる植物は、たとえ同じ植物種でも、異なる資源であることが予想されます。では、植物の性差はどのような影響を昆虫に与えるのでしょうか? この疑問は昆虫の植物利用の進化を考える上でとても興味深いものと思われますが、これまでに、答えとなるべき例は知られていませんでした。
ヒサカキの雌木で見つからないソトシロオビナミシャク

写真1: (a) ヒサカキ、(b) ソトシロオビナミシャクの終齢幼虫がヒサカキの雄花蕾を食害している様子、(c) ソトシロオビナミシャクの成虫
(クリックで拡大します)
ヒサカキは常緑の中低木で(写真1a)、雄花(葯のみがある花)のみをつける雄木、雌花(胚珠のみがある花)をつける雌木、両性花(葯と胚珠がある花)をつける両性木があります。但し、両性木には、両性花のみをつける木だけでなく、雄花と両性花、雌花と両性花、雄花と雌花と両性花をつける木まであります。このヒサカキは様々な昆虫に利用されていますが(Tsuji & Sota 2013)、観察していると、雄木や両性木では見つかるのに、雌木では見つからない幼虫がいるのです(写真1b)。本当にこのような現象があるのかと、信じられなかったので、山の一斜面に生えているヒサカキの花を片っ端から観察しましたが、やはり、雄木と両性木では見つかるのに、雌木では見つかりませんでした。この幼虫はソトシロオビナミシャク(以下ナミシャク)(写真1c)という花だけを食べるシャクガで、春はツツジの花蕾を冬はヒサカキの花蕾を食べています。とても面白い材料と考え研究をすすめることにしました。
ナミシャクは雌花を食べると死ぬ

図1: 様々な部位の雄または雌の花蕾を食べたときの、ナミシャクが成虫になるまでの発育日数。アルファベットが異なるものは発育日数が有意に異なることを表します。雌の萼と花弁、胚珠を食べたときは幼虫が全て死亡したため発育日数は存在していません。
(クリックで拡大します)
まず、卵から生まれたばかりの幼虫に、雄または雌のヒサカキの枝(花蕾と葉がついている)を提示して、食べるかどうか調べてみました。すると、雄のヒサカキを与えられた幼虫は花蕾のみを食べ、成長を続ける一方、雌花を与えられた幼虫は数個の花蕾のみを食べた後に、ほぼ全ての幼虫が死亡しました(図1)。この結果から、雌木で幼虫が生きられないから、野外の雌ヒサカキで幼虫が見つからなかった、ということが分かりました。
そもそも、雄花と雌花では、葯、胚珠と存在する器官が違うので、その違いが幼虫の生存を決めている可能性があります。そこで、どの花の器官が幼虫の生存を左右するのかを調べるため、幼虫の摂食する花の器官を操作する実験を行いました。ナミシャクが食べる蕾はまだ硬く、完全に萼で覆われています。そのため、何の操作もせず、そのまま花蕾を与えると、幼虫は、まず萼を食べて穴をあけ、その後に花弁、葯もしくは胚珠を食べます。そこで、幼虫が直接花弁などを食べられるよう、蕾を半分に切り与えてみました。すると、幼虫は萼を食べず、花弁と葯・胚珠を食べました。さらに、幼虫が花弁だけを食べる状況を作るため、蕾を半分に切り、葯や胚珠を取り除いて与えました。その結果、雌花であっても、胚珠と花弁、もしくは花弁のみを食べれば、幼虫は無事に成長することが分かりました(図1)。つまり、幼虫は雌花の萼を食べた場合にのみ、死んでしまったのです。予想とは裏腹に、雄花と雌花の胚珠と葯という異なる器官ではなく、共通する器官である萼の雌雄差が幼虫の生存を左右していました。
この飼育実験の結果から、萼の化学防御物質に雌雄差があると考え、防御物質の代表ともいえる縮合タンニン量・総フェノール量の含有量を測定したところ、これらの防御物質は雌花の萼では雄花の2倍程度も含まれていることが分かりました。
母親が雄花を選んで産卵する行動の進化
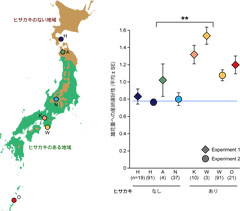
図2: ナミシャクの産卵選好性。グラフの水色の線は雄花と雌花に同数の卵を産卵したことを示し、線より上にいくほど、雄花により多く産卵したことを示しています。地図上にナミシャクの採集地点を示し、それぞれの色はグラフの結果と対応しています。Tsuji and Sota (2011)を改変。
(クリックで拡大します)
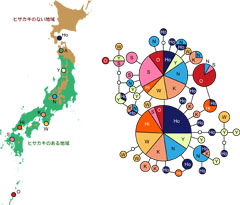
図3: ソトシロオビナミシャクのCOI領域の830塩基をもとに作成したハプロタイプネットワーク。ネットワークのそれぞれの丸は一つのハプロタイプを示し、丸が大きいほど、その塩基配列を持つ個体が多かったことを示します。線でつながった丸の間では、調べた830塩基のうち1塩基だけが違うことを示します。小さい白丸は、サンプルからは見つからなかった塩基配列です。採集地点ごとに色分けを行いました。地図上でも丸の大きさは個体数を示します。Tsuji and Sota (2011)を改変。
(クリックで拡大します)
幼虫が食べると死んでしまう雌花に、成虫は産卵するのでしょうか? これを調べるため、京都の瓜生山でライトトラップによる夜間採集を行い、ナミシャクを採取し、産卵選好性の実験を行いました。すると、このナミシャクは、雌木ではなく、雄木の花蕾周辺によく産卵することが分かりました。この適応的な産卵行動は、ヒサカキへの適応として進化したのか?を明らかにしたいと考え、さらなる実験を行うことにしました。このナミシャクは北海道以南に生息しますが、ヒサカキは東北以南の低地にしか分布していません。そこで、ヒサカキの分布していない地域のナミシャクと、ヒサカキが分布している地域のナミシャクを採集し、産卵行動を比較しました。すると、ヒサカキの分布しない地域に比べ、分布している地域では有意に雄花に対する産卵選好性が強いことが分かりました(図2)。
そこで、ヒサカキの分布する地域と分布していない地域のナミシャクは遺伝的に分化しているのではと考え、ミトコンドリアDNAのシトクロムオキシダーゼI(COI)と呼ばれる領域の塩基配列830塩基を読み、解析を行いました。その結果、206個体の塩基配列が得られ、地理的にほとんど遺伝的に分化していないことが明らかになりました(図3)。図3で示されているハプロタイプとは、塩基配列のタイプのことで、ネットワークは各ハプロタイプ間の関係を示しています。色分けは地図上の採集地点と対応しています。この図で、全採集地点のナミシャクが大きな2つの丸に含まれていることから、北海道から九州、沖永良部島に至るまで、共通のハプロタイプを持っていることが分かります(これまでは、このナミシャクの分布域は屋久島までとされていましたが、トカラ海峡を隔てた沖永良部島に分布していることも調査の中で新たにわかりました)。遺伝分化がほとんどないことから、ごく最近に分布拡大した、もしくはナミシャクが非常に大きな分散能力を持っていることが示唆されます。
これらの産卵実験・遺伝解析から、ナミシャクの適応的な産卵行動は、ヒサカキを利用するうえで進化した、それも、COI遺伝子では分化が検出できないくらいに急速に進化を遂げていることがわかりました。
おわりに
植物の性差により、幼虫の生死が左右され、しかも母親は、まだ硬い蕾の時期に植物の性を区別し産卵する行動を進化させている、という初めて見つかった現象について紹介しました。鱗翅目では、数百年という短い時間で好んで産卵する植物を変化させる行動の進化が知られているため、今回のシャクガが急速に雄と雌の植物を選び分ける行動を進化させていたとしても不思議ではありません。今後、ヒサカキの花蕾にはどのような雌雄差があり、成虫がどのようにヒサカキの性を認識しているのか、また、ヒサカキの花を利用する他の昆虫はどのように雌雄の木を利用しているのか、を調べることで、さらにおもしろいことが分かるのではないかと期待しています。
参考文献
- Tsuji K, Sota T. (2010) Sexual differences in flower defense and correlated male-biased florivory in a plant-florivore system. Oikos 119: 1848-1853.
- Tsuji K, Sota T. (2011) Geographic variation in oviposition preference for male and female host plants in a geometrid moth: implications for evolution of host choice. Entomol. Exp. Appl. 141: 178-184.
- Tsuji K, Sota T. (2013) Florivores on the dioecious shrub Eurya japonica and the preferences and performances of two polyphagous geometrid moths on male and female plants. Entomol. Sci. 16: 291-297.
« シャーレの中での実験から野外での保全を考える | コラム一覧 | ベニシジミのメスのセクシャルハラスメント回避行動 »
応用動物学/応用昆虫学コラム
応用動物学/応用昆虫学の分野でいま注目されている研究成果を、第一線で活躍している研究者が解説します。
日本応用動物昆虫学会(応動昆)
「むしむしコラム・おーどーこん」は、日本応用動物昆虫学会電子広報委員会が管理・運営しています。