2013年01月14日掲載 【オオヒラタシデムシにおける飛翔能力の退化過程】
飛翔能力の獲得は、昆虫の長い進化史の中でも特に重要なイベントの一つです。しかし、飛ぶには多くのエネルギーが必要なため、このような飛翔能力が退化して飛べなくなった種も多くいます。今回は、種内で飛翔能力に2型の見られる種、オオヒラタシデムシにおける飛翔能力の退化過程について調べた研究を紹介します。
はじめに
昆虫は非常に種数の多い分類群として知られています。約4億年前に起きたとされる飛翔能力の獲得は、この莫大な種数をもたらした要因の一つだと考えられています。飛翔能力というのは、長い距離を移動できるとともに、餌や交配相手を探すうえでも有利な能力です。しかし、飛ぶための器官を持つには多くのエネルギーが必要とされるため、様々な分類群において飛翔能力は退化してきました。そして、退化の途中段階にあると考えられる、飛翔能力に多型を持つ種も多く知られています。飛翔能力に多型がある種というのは、飛翔能力がどのような過程を経て退化してきたかを考える上で、とても興味深い研究対象です。
オオヒラタシデムシにおける飛翔筋2型の発見

図1: (a)オオヒラタシデムシ。(b)飛翔筋のある個体を解剖して間接飛翔筋を出したところ。矢印で示したものが筋肉。(c)飛翔筋のない個体。代わりに別の組織が筋肉のあった部分を占めています。
(クリックで拡大します)
これまでの研究から、甲虫目シデムシ科のオオヒラタシデムシ(Necrophila japonica; 図1a)でも、飛ぶための筋肉である飛翔筋に2型があることが分かりました(図1b, c)。オオヒラタシデムシは日本に広く分布する種で、公園などでも普通に見つけることができるため、見たことのある方も多いかもしれません。私は、この種において、飛翔筋のある個体とない個体の地理的な分布と、遺伝分化のパターンを調べることで、飛翔能力の退化過程とその要因を明らかにできないかと考え、研究を行いました。
日本における遺伝分化パターンと、飛べる個体、飛べない個体の分布パターン
私は日本全国でオオヒラタシデムシを採集し、ミトコンドリアDNAのシトクロムオキシダーゼI(COI)と呼ばれる領域の塩基配列842塩基を読んで、日本全体での遺伝的な分化のパターンを調べました。この領域は進化の速度が比較的早いため、種内の集団の間での分化パターンを見るのに良く使われます。そして、飛翔筋のあるなしのパターンも日本全国で調べ、これらの間に関係があるかどうかについて検討してみました。
遺伝子解析の結果からは、396個体から塩基配列が得られ、遺伝的な分化の程度は大きくはないものの、地理的にある程度分化している傾向が明らかになりました(図2)。図2で示されているハプロタイプとは、塩基配列のタイプのことで、ハプロタイプネットワークとは、各ハプロタイプ間の関係を示したものです。塩基配列の近いハプロタイプは同じ色で示してあります。そして、図2の地図上には、色で分けたハプロタイプの各グループの分布が示されています。北海道の個体は全て青のグループの塩基配列を持ち、東北地方の個体は全て水色、関東も大半が水色になりました。それに対して関東よりも西側の地域ではオレンジを中心により多様な塩基配列を持つ個体が分布している傾向がみられました。遺伝分化の程度があまり大きくないことから、比較的最近の分布拡大によって現在の分布が形作られたようです。

図2: オオヒラタシデムシのCOI領域の842塩基をもとにして作成したハプロタイプネットワークと、各ハプロタイプのグループの地理的な分布。ハプロタイプネットワーク上のそれぞれの丸は一つのハプロタイプを示しており、丸が大きいほど、その塩基配列を持つ個体が多かったことを示します。線でつながった丸の間では、調べた842塩基の塩基配列のうち1塩基だけ配列が違うことを示します。小さい黒丸は、サンプルからは見つからなかった塩基配列です。ハプロタイプネットワーク上で塩基配列の近いものを同じ色でまとめてあります。地図上でも丸の大きさは個体数を示します。Ikeda and Sota (2011)を改変。
(クリックで拡大します)
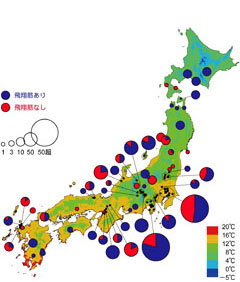
図3: 飛翔筋のある個体及びない個体の地理分布。丸が大きいほど、サンプル数が多いことを示します。地図上の色は年平均気温。Ikeda and Sota (2011)を改変。
(クリックで拡大します)
次に、オオヒラタシデムシを解剖して飛翔筋の有無を調べ、それが日本全体でどのように分布するかを調べました。雌242個体、雄218個体について調べたところ、飛翔筋のある個体とない個体の割合は雌と雄の間で違いがなく、ともに70%ぐらいの個体が飛翔筋を持っていました。地理的な分布をみたところ、日本全国に飛べる個体も飛べない個体も広く分布しており、飛翔筋の有無には地理的にはっきりとしたパターンというのはみられませんでした(図3)。ですが、北海道の中部や東北地方の北部で採集された個体は全て飛翔筋を持っていました。
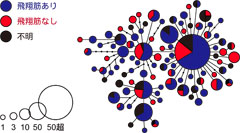
図4: ハプロタイプネットワークと飛翔筋の有無の関係。Ikeda and Sota (2011)を改変。
(クリックで拡大します)
そして、これらの結果をもとに、飛翔筋の有無と遺伝分化パターンの地理的な関係について検討するため、ハプロタイプネットワーク上に飛翔筋の有無のデータをのせて関係を調べたところ、これらの間には関係は認められませんでした(図4)。多くのハプロタイプで飛べる個体も飛べない個体も存在し、ハプロタイプの近い個体間でも飛翔能力には特に似たようなパターンはありませんでした。これらのことから、飛翔能力の退化が系統的にどこで生じたかにははっきりとした傾向はないようで、大きな地理スケールで見ると、飛翔筋2型の状態が保たれながら日本の様々な地域に分布を広げ、集団間で遺伝的に分化してきたようです。
飛翔能力と生息環境の関係
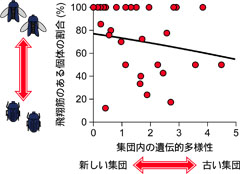
図5: 集団内の遺伝的多様性と飛翔筋のある個体の割合の関係。Ikeda and Sota (2011)を改変。
(クリックで拡大します)
これまでの解析からは、オオヒラタシデムシにおける飛翔能力の退化にははっきりとした地理的パターンや、ハプロタイプネットワーク内で見られるような系統的な関係は認められませんでした。それでは、飛翔能力と生息環境はどのように関係しているのでしょうか。一般には、短期間で消失するような生息地や、新しくできた生息地には、飛べる個体の方がより侵入しやすい傾向があります。そのような環境のもとで形成された歴史の浅い集団では、少ない個体が侵入して個体数を急速に増やすため、遺伝的に似た個体が多くなり、遺伝的な多様性は低くなります。したがって、遺伝的多様性を集団の新しさの指標と考えると、遺伝的多様性の低い新しい集団ほど、飛べる個体の割合は高いという関係がみられることが予想されます。これについて調べてみると、予想した通り、遺伝的多様性の低い集団、つまり新しいと考えられる集団ほど、飛翔筋のある個体の割合が高いという結果が得られました(図5)。
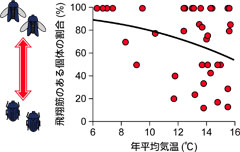
図6: 年平均気温と飛翔筋のある個体の割合の関係。Ikeda and Sota (2011)を改変。
(クリックで拡大します)
次に、新しい集団はどのような地域に形成されてきたのかを考えてみました。遺伝子解析の結果では、オオヒラタシデムシは比較的最近分布を拡大し、また、東北地方などの寒い地域では集団間の分化の程度は小さい傾向がみられました。このようなパターンは他の生物でも知られており、約20,000年前の最終氷期が終わった後の気温上昇に伴って、急速に寒冷な地域に分布を拡大したためだと考えられています。遺伝分化パターンからは、オオヒラタシデムシでもそのような分布拡大が生じたのではないかと予想されます。そこで、気温と飛翔筋の有無の割合の関係を調べることで、この仮説を検証してみました。急速な分布拡大は主に飛べる個体の移動によると考えられるため、寒い地域に急速に分布を拡大したのであれば、気温が低い地域ほど飛べる個体の割合が高いと予想されます。この関係を調べたところ、この仮説を支持するようなパターンがみられました(図6)。これらのことから、飛べる個体は、寒い地域に新しく形成された集団で多いというパターンが得られました。地図上でも、より寒い東北地方や北海道の中部では飛べる個体の割合が高い傾向がみられており(図3)、東北地方と関東地方のハプロタイプのグループが同じで、これらの地域の間であまり遺伝的に分化していない(図2)ことを考えると、東北地方の集団というのは最終氷期後に関東地方からの急速な分布拡大によって形成されてきたと考えられます。
これに対してより暖かい地域に存在する集団というのは、気候変動の中でも維持されてきた、より長期的に存在してきた集団だと考えられます。一般に飛翔能力というのは、飛ぶ必要のないような、長く維持されている安定した環境で退化しやすいと言われています。今回の結果でも暖かい地域の古い集団の方が飛べない個体は多い傾向がみられており、もしかすると、このように長く維持されている暖かい地域において、オオヒラタシデムシの飛翔能力は今後退化していくのかもしれません。
おわりに
ここまで、オオヒラタシデムシの飛翔筋2型について明らかになったことを紹介してきました。一般に、飛ぶ個体と飛ばない個体の間では、産卵などの繁殖に使うエネルギー量に違いがあり、飛ばない個体の方が、より多くのエネルギーを繁殖に回すことが知られています。シデムシで種間で比較した場合にはこのような飛翔能力と繁殖投資の明確な関係性が明らかになっていますが(Ikeda et al. 2008)、オオヒラタシデムシの種内の2型間ではそのような傾向は見つかっていません。飛べない個体では筋肉がない分のエネルギーを何か別のことに使用していると考えられるのですが、まだそれが何なのかはわかっていません。オオヒラタシデムシは、現在、広い地域で飛べる個体も飛べない個体もみられていますが、なぜこの状態が維持されているのか、そして今後どのような場合に退化する可能性があるのかについて、2型間での違いを明らかにすることで、さらに何か見えてくるのではないかと思います。飛翔多型種における飛翔能力以外の違いを明らかにすることで、どのような場合に飛翔能力を持たないことが有利になり、そして退化していくのか、を明らかにするための重要な手掛かりが得られると考えられます。このような研究を続けていくことで、昆虫における飛翔能力の退化に関して、今後さらに様々な知見が明らかになるでしょう。
参考文献
- 池田紘士 (2011) 飛べないシデムシ―その生態と系統地理―. 昆蟲(ニューシリーズ) 14: 133-138.
- Ikeda H, Kagaya T, Kubota K, Abe T. (2008) Evolutionary relationships among food habit, loss of flight, and reproductive traits: Life history evolution in the Silphinae (Coleoptera: Silphidae). Evolution 62: 2065-2079.
- Ikeda H, Kubota K, Kagaya T, Abe T. (2007) Flight capability and food habits of silphine beetles: are flightless species really "carrion beetles"? Ecol. Res. 22: 237-241.
- Ikeda H, Sota T. (2011) Macro-scale evolutionary patterns of flight muscle dimorphism in the carrion beetle Necrophila japonica. Ecol. Evol. 1: 97-105.
« 一見すると不合理な、テントウムシのエサ選び | コラム一覧 | 研究室紹介: 京都大学大学院理学研究科・動物生態学研究室 »
応用動物学/応用昆虫学コラム
応用動物学/応用昆虫学の分野でいま注目されている研究成果を、第一線で活躍している研究者が解説します。
日本応用動物昆虫学会(応動昆)
「むしむしコラム・おーどーこん」は、日本応用動物昆虫学会電子広報委員会が管理・運営しています。