2012年12月14日掲載 【偶然か?必然か? レトロポゾン挿入が関与したと思われるガ類種分化の例】
ガ類の多くは、メスが種ごとに異なる性フェロモンを分泌し同種オスを呼び寄せて交尾します。性フェロモン成分が種ごとに違うのは近縁種同士が間違って交尾することを防ぐためです(生殖前隔離)。故に、性フェロモン成分の生産は、性質の異なる酵素群によって厳密に制御されています。しかし、我々はウスジロキノメイガという蛾において、本種が近縁種ではレトロポゾンのゲノムへの挿入により不活化してしまった酵素遺伝子を用いて、近縁種とは似て非なるフェロモンを生産していることを明らかとしました。今回はガ類の種分化に影響するフェロモンの作り分けの「なぜ?」を紹介します。
昆虫が地球上で確認されている生物種の75%以上を占めている事は、昆虫が環境に適応する為の生存戦略を獲得し、繁栄に成功した結果だと考えられています。例えば、匂いや、音、光を用いた個体間のコミュニケーションは、昆虫達が広大な空間から有利に配偶者を見つけ交配する為に獲得した生存戦略の一つです。中でもガ類で研究が進められている性フェロモンを介した匂いのコミュニケーションはメスが同種のオスを選択的に誘引し、その結果、交雑種の出現を防いでいる為、生殖前隔離の役割を担っています。その為、近縁種を正しく認識する為に、ガ類性フェロモン成分とその比率は、複数のフェロモン生合成酵素の協奏的な働きにより厳密に支配されている必要があります。その酵素群の一つに、ガ類のフェロモン腺で発現する11位不飽和化酵素(官能基側から数えて11番目と12番目の炭素間に二重結合を挿入する反応を触媒する酵素。本文中では以下Δ11とする)が知られています。
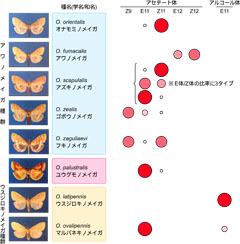
図1: 日本産アワノメイガ類(Ostrinia)の性フェロモン分子の多様性(Ishikawa et al. 1999を改変)。
国内に生息するアワノメイガ類の形態は類似しているが性フェロモン成分は種毎に異なる。アワノメイガ類の性フェロモン分子は炭素鎖数14のアセテートとアルコールが基本骨格であり、赤い円は
フェロモン成分の二重結合の位置と向き(例E11→11位にトランス体の二重結合を含む)を示し、円の面積はそれら成分のおおまかな比率を意味する。種群(青色: アワノメイガ種群、桃色: ユウグモノメイガ、黄色: ウスジロキノメイガ種群)は外部形態の相違とミトコンドリアCOII遺伝子の配列を元にグループ化した。
(クリックで拡大します)
ところで、ガ類の仲間には農業害虫が多く含まれますがアワノメイガ(Ostrinia furnacalis)もその一つであり、トウモロコシの大害虫として良く知られています。国内に生息するアワノメイガ属昆虫はΔ11が産み出すフェロモン成分の比率と種認識を考えるのに好適な材料です。国内産アワノメイガはアワノメイガ種群、ユウグモノメイガ、ウスジロキノメイガ種群に分類され、性フェロモン成分は炭素鎖が14の不飽和脂肪族のアセテートかアルコールに限られています(図1: Ishikawa et al. 1999)。表にして比べてみると、国内産アワノメイガの性フェロモン識別には二重結合のトランス体(E体)とシス体(Z体)の混合比に何か重要な意味がありそうです。アワノメイガ種群では、不飽和化酵素のアミノ酸配列が相同でE体とZ体を決まった比率で同時に生産し、種ごとの比率の制御は別の酵素が決定することが分かっています(Fukuzawa et al. 2006; Lassence et al. 2010)。それではE体の性フェロモンしか分泌しないウスジロキノメイガ種群でも、同じようにE体とZ体の両方の原料からE体の性フェロモンのみを選択して作っているのでしょうか?
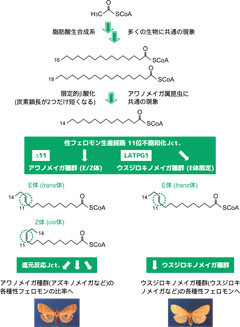
図2: アワノメイガ属昆虫の一般的なメス性フェロモン生合成経路における幾何異性体の作り分け。
アワノメイガ種群では還元酵素でE/Z体の異性体比が制御される一方、ウスジロキノメイガ種群は不飽和化酵素LATPG1がE体のみ産生する反応を触媒する。
(クリックで拡大します)
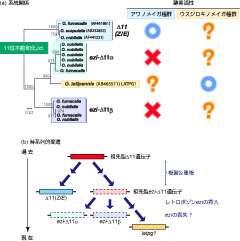
図3: アワノメイガ属の性フェロモン生合成に関与する11位不飽和化酵素のa)系統関係とb)時系列的変遷。
(a) ウスジロキノメイガ種群の性フェロモン生合成に関与するLATPG1はアワノメイガ種群でレトロポゾンの挿入により不活性化されたグループに含まれた(Fujii et al. 2011)。
(b) 現在, ウスジロキノメイガ種群でLATPG1が活性を有する理由として、時間の変遷の中で一度ezi-が挿入したコピーからeziが喪失してlatpg1が活性化した為とする説がある(Rooney et al. 2011)。
(クリックで拡大します)
この疑問をひもとく為に、我々はウスジロキノメイガ(Ostrinia latipennis)において性フェロモン腺で発現するΔ11遺伝子latpg1をクローニングして機能を明らかにし、このΔ11がE体のフェロモン原料のみを生産する事を示しました(図2: Fujii et al. 2011)。そして興味深い事に、アワノメイガ種群のゲノム上でlatpg1と相同な遺伝子が存在しているにも関わらず、eziと呼ばれるレトロポゾンのイントロンへの挿入により転写が不活化していることが知られていました(図3a: Xue et al. 2007; Rooney 2011)。この事実の説明として、交配が繰り返される過程でウスジロキノメイガのゲノムからeziの配列が抜け、その結果latpg1が機能する事が出来るようになったと考えています(図3b: Fujii et al. 2011; Rooney et al. 2011)。
この可能性を検証する為に、現在はウスジロキノメイガのゲノムを広範に調査しています。これによりウスジロキノメイガのゲノム上からもアワノメイガ種群のΔ11の類似遺伝子を発見することが出来るかもしれません。

図4: トランスポゾンとレトロポゾンの違い。
トランスポゾンはゲノム上から切り出されてDNAとして転位するのに対し、レトロポゾンはDNAから一旦RNAを合成し、自身の逆転写酵素(RT)によりcDNAとしてゲノム上に組み込まれる点が異なる。 結果的にレトロポゾンのこの性質は不飽和化酵素遺伝子の多コピー化の原因となりうる。
(クリックで拡大します)
ガ類昆虫の11位不飽和化酵素は祖先型から重複と複製を繰り返し多コピー化したと言われます(図3b: Rooney et al. 2011)。その中でレトロポゾンの挿入は間違いなくこの多コピー化に貢献していたはずです(図4)。この多コピー化の過程で起こった酵素の多様化と種の分化を考察する為には更なる研究が必要ですが、少なくとも生殖前隔離に関わる重要な機能を司るフェロモン生合成酵素遺伝子にezi挿入というイベントが起こり、性フェロモン成分の変更が種分化に何らかの影響を与えたと考えられます。
参考文献
- Fujii T, Ito K, Tatematsu M, Shimada T, Katsuma S, Ishikawa Y. (2011) A sex pheromone desaturase functioning in a primitive Ostrinia moth is cryptically conserved in congeners' genomes. Proc. Natl. Acad. Sci. U.S.A. 108(17): 7102-7106
- Fukuzawa M, Fu X, Tatsuki S, Ishikawa Y. (2006) cDNA cloning and in situ hybridization of Δ11-desaturase, a key enzyme of pheromone biosynthesis in Ostrinia scapulalis. J. Insect Physiol. 52(5): 430-435.
- Ishikawa Y, Takanashi T, Kim C, Hoshizaki S, Tatsuki S, Huang Y. (1999) Ostrinia spp. in Japan: their host plants and sex pheromones. Entomol. Exp. Appl. 91: 237-244.
- Lassance JM, Groot AT, Lienard MA, Antony B, Borgwardt C, Andersson F, Hedenstrom E, Heckel DG, Lofstedt C. (2010) Allelic variation in a fatty-acyl reductase gene causes divergence in moth sex pheromones. Nature 466: 486-489.
- Rooney AP. (2011) Pheromone emergencies and drifting moth genomes. Proc. Natl. Acad. Sci. U.S.A. 108(20): 8069-8070.
- Xue B, Rooney AP, Kajikawa M, Okada N, Roelofs WL. (2007) Novel sex pheromone desaturases in the genomes of corn borers generated through gene duplication and retroposon fusion. Proc. Natl. Acad. Sci. U.S.A. 104(11): 4467-4472.
応用動物学/応用昆虫学コラム
応用動物学/応用昆虫学の分野でいま注目されている研究成果を、第一線で活躍している研究者が解説します。
日本応用動物昆虫学会(応動昆)
「むしむしコラム・おーどーこん」は、日本応用動物昆虫学会電子広報委員会が管理・運営しています。