2014年01月01日掲載 【共生界の遺伝的フランケンシュタインモンスター】
農業害虫として悪名の高いミカンコナカイガラムシは、別種の細菌を内部に宿した細菌がさらに昆虫の細胞の中に入り込むマトリョーシカ人形のような入れ子状共生系を持っています。この共生系の進化には、これら3種の以外の生物も深く関わって来たことがゲノム解析により明らかになりました。
入れ子か否か



図1: コナカイガラムシと入れ子状共生系。(A)ミカンコナカイガラムシ、(B)腹部の菌細胞塊(赤)、(C)入れ子状共生系、ベータ細菌(赤)の中にガンマ細菌(緑)が入り込んでいる。青色は宿主細胞の核。画面中央で2個の菌細胞が隣接している。
(クリックで拡大します)
綿ぼこりのようなふわふわで小さく扁平な体を覆い、動きは非常に緩慢な虫が植物の表面に集団を作っているのを見たことはないでしょうか? おそらく彼らはコナカイガラムシです(図1)。農業害虫が多く含まれるこの科の昆虫は専ら寄主植物の汁を吸って生きているのですが、植物の汁は栄養的に非常にアンバランスで、普通であれば動物はこのような食物だけでは生きて行けません。しかし、コナカイガラムシはそんなことお構いなしに爆発的に増えることができます。それは植物の汁にほとんど含まれていない必須アミノ酸などの不可欠な栄養素を合成し、昆虫に与えてくれる必須細胞内共生細菌を保有しているからです。このコナカイガラムシの共生細菌は宿主の系統によって異なった特徴を持っています。コナカイガラムシ科はコナカイガラムシ亜科とワタカイガラモドキ亜科の二つに分かれるのですが、前者に属する多くの種はトレンブレイヤ(Tremblaya、以下ベータ細菌)とモラネラ(Moranella、以下ガンマ細菌)の2種類の必須共生細菌を、 後者に属する種の多くはベータ 細菌のみを保有しています。どうも二つの亜科に分岐したあとに、コナカイガラムシ亜科の方でガンマ細菌を獲得したようです。どちらの場合においても、共生細菌は細菌との共生に特殊化した菌細胞という細胞の細胞質に局在しています(図1BC)。面白いことに、コナカイガラムシ亜科において先の二種の共生細菌は菌細胞内で単に共存しているのではなく、ガンマ細菌がベータ細菌の中に入り込んでいるのです。つまり、コナカイガラムシの細胞の中にベータ細菌がおり、さらにこのベータ細菌の中にガンマ細菌が入っている、例えるならロシアのマトリョーシカ人形のような入れ子状の共生関係を構築しているのです(図1C)(Dohlen et al. 2001)。
近年、ミカンコナカイガラムシ(Planococcus citri、以下ミカンコナ)の共生細菌のゲノム配列が決定され、それぞれの細菌の持っている遺伝子が相互に組み合わされて初めて宿主昆虫に取って必須な機能を発揮しうることが示唆されました。一方、ワタカイガラモドキ亜科の共生細菌については微生物学的な解析しかなされていませんでしたが、二つの亜科のカイガラムシが同一の寄主植物で生育可能な例があることから(Koga et al. 2013)、コナカイガラムシ亜科では2種の共生細菌で果たしている機能をワタカイガラモドキ亜科のベータ細菌は単独で担いうることが考えられました。そのため、ガンマ細菌の獲得によって二つの亜科のベータ細菌のゲノム間にどのような違いが生じたのか、どのような遺伝子レパートリーで2種の細菌と同一の機能を果たしているのか、など比較ゲノム的な興味がわきました。そこで、ワタカイガラモドキ亜科に属するキュウコンコナカイガラムシ(Phenacoccus avenae、以下キュウコンコナ)のベータ細菌のゲノムを決定し、ゲノム構造や推定代謝経路の比較をおこなうことにしました(Husnik et al. 2013)。
ミカンコナのベータ細菌のゲノムはガンマ細菌の獲得によってさらに縮退した
キュウコンコナのベータ細菌のゲノムサイズは170 kbで、175個の遺伝子をコードしており、全長140 kbで121個の遺伝子をコードしているミカンコナのベータ細菌のゲノムの大半に加えて、ミカンコナのベータ細菌では失われていた翻訳関係の遺伝子なども保持していました。またゲノム上の遺伝子の密度も高くなっていました。これらの結果は、ミカンコナの祖先がガンマ細菌を獲得することでベータ細菌のゲノムへの選択圧が減少したため、多くの遺伝子が欠失し、ミカンコナのベータ細菌ゲノムはキュウコンコナのベータ細菌に比べてさらに縮退したことを示しています。
キュウコンコナのベータ細菌は単独でミカンコナの二種の共生細菌の機能を果たす
これら共生細菌の担う生物機能の一つは必須アミノ酸の合成ですが、その合成経路に関わる遺伝子をミカンコナの複合共生系とキュウコンコナのベータ細菌との間で比較したところ、 ミカンコナではベータとガンマ細菌の遺伝子が組み合わさって構成されている合成経路をキュウコンコナのベータ細菌は単独で構成していることが判明しました。この結果からキュウコンコナの共生系とミカンコナの複合共生系が同じ生物学的機能を担っていることが予想されました。
ただ、これら二種のコナカイガラムシの共生系において、合成経路の全ての遺伝子が揃っているのは十種類の必須アミノ酸のうちアルギニンとトリプトファンの二つのみであり、かなりの数の必要な遺伝子が欠失していたのです。これらの欠失した遺伝子がどのようにして補われているのかを明らかにするために、共生細菌が格納されている菌細胞において発現が亢進している遺伝子をミカンコナについて探索しました。
宿主や他の細菌が埋めていた代謝経路の穴
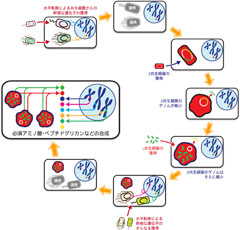
図2: ミカンコナカイガラムシの内部共生システムの進化過程
(クリックで拡大します)
その結果、ほとんどの欠失遺伝子について類似の機能を果たす遺伝子の発現が亢進していました。しかも、これらの遺伝子の配列を解析したところ、そのいくつかは昆虫ではなく細菌、それもベータでもガンマ細菌とは全く異なるボルバキアやリケッチア、カルディニウムなど昆虫類に広く見られる共生細菌の遺伝子と高い類似性を示したのです。このような細菌由来の遺伝子は必須アミノ酸合成系に限らず、ビタミンや細胞壁合成経路にも関与しており、最低でも 22個が菌細胞で発現していることが確認されました。さらに宿主昆虫のゲノム解析の結果、全てミカンコナの核ゲノム上に存在することが明らかになりました。つまり、これらの遺伝子はミカンコナの祖先に感染していた共生細菌から宿主昆虫の核ゲノムに水平転移したものだったのです。現時点でミカンコナは水平転移遺伝子の元となった共生細菌に感染していないことを考えると、これらの共生細菌は遺伝子の一部をミカンコナの核ゲノムに転移したあとに、その感染が失われたようです(図2)。
昆虫と(少なくとも)5種の細菌が織りなすモザイク共生系
以上の結果から、ガンマ細菌がベータ細菌の中に入り込み、これがさらに宿主の細胞内に入り込むという複雑な入れ子状の共生系をもつミカンコナの代謝系は、宿主昆虫や2種の共生細菌に加えて、3種を下らない(おそらく6種以上)細菌から水平転移によって獲得したさまざまな遺伝子が、パッチワークのように組み合わさって構築される複雑なものであることが判明しました。この発見は、昆虫と細胞内共生細菌の関係にとどまらず、オルガネラとは何なのか、宿主とオルガネラとの関係がどのように構築され、細胞内共生が進化するのか、といった問いに新たな観点を与えるものであります。
今後の展開
今回の成果によって共生系を構築する複数の生物間で遺伝子産物もしくはその生成物がやり取りされていることが示唆されましたが、具体的にどのような分子メカニズムで実行され、この複雑な共生系が維持されているのかを明らかにする研究が重要であると考えています。
引用文献
- von Dohlen CD, Kohler S, Alsop ST, McManus WR. (2001) Mealybug β-proteobacterial endosymbionts contain γ-proteobacterial symbionts. Nature 412:433-436. doi:10.1038/35086563
- Husnik F, Nikoh N, Koga R, Ross L, Duncan RP, Fujie M, Tanaka M, Satoh N, Bachtrog D, Wilson ACC, von Dohlen CD, Fukatsu T, McCutcheon JP. (2013) Horizontal Gene Transfer from Diverse Bacteria to an Insect Genome Enables a Tripartite Nested Mealybug Symbiosis. Cell 153: 1567-1578. doi:10.1016/j.cell.2013.05.040
- Koga R, Nikoh N, Matsuura Y, Meng X-Y, Fukatsu T. (2013) Mealybugs with distinct endosymbiotic systems living on the same host plant. FEMS Microbiol. Ecol. 83: 93-100. doi:10.1111/j.1574-6941.2012.01450.x
« 「生物を生きたまま電子顕微鏡で高解像度観察する」~昆虫が分泌する物質を規範とした"防護服"ナノスーツの開発~ | トピック一覧 | イモムシ体表にスポット紋様が生じるメカニズムの解明 »
応用動物学/昆虫学最新トピック
プロの研究者でもまだ知らないような、出来たてホヤホヤの最新研究成果を分かりやすくお伝えします。
日本応用動物昆虫学会(応動昆)
「むしむしコラム・おーどーこん」は、日本応用動物昆虫学会電子広報委員会が管理・運営しています。