2014年02月10日掲載 【オルガネラ様共生細菌で身を守るミカンキジラミ】
昆虫と微生物の共生のかたちのひとつに、微生物が宿主を外敵から守る「防衛共生」がありますが、この関係は進化的に不安定と考えられてきました。ところがさきごろ、私たちは、カンキツ類の大害虫であるミカンキジラミが、毒を作る共生細菌を自らの一部として安定な関係を築き、これを武器に天敵から身を守っているらしいことを明らかにしました。
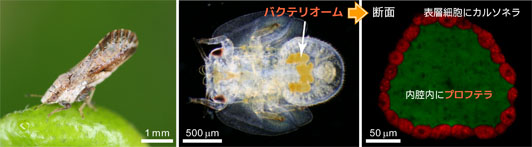
図1: (左) ゲッキツ上のミカンキジラミ成虫。(中) ミカンキジラミ5齢幼虫。腹部に見える黄色いクロワッサン型の構造が、共生器官「バクテリオーム」。(右) バクテリオームの断面。表層細胞内にカルソネラ(赤色)、内腔内にプロフテラ(緑色)が多数収納されている。
(クリックで拡大します)
昆虫と共生する微生物は、その機能にもとづき、宿主のえさに乏しい栄養分を供給する「栄養共生体」、毒を合成して宿主を天敵から守る「防衛共生体」などに分けることが出来ます。宿主の栄養要求は大きく変動しないことから、栄養共生体の多くは進化的に安定な地位にあり、宿主の生存に不可欠で、あらゆる個体に存在し、特別な「共生器官」に収納され、垂直感染のみにより虫の親から子へと永続的に受継がれ、宿主と共種分化し、その過程でゲノムサイズが大幅に縮小(<100万塩基対)するなど、ミトコンドリアや葉緑体といったオルガネラと同様、宿主と融合・一体化する傾向を示します。これに対して、外敵にまつわる環境要因はめまぐるしく変動するうえに、毒を作る共生体を維持するためのコストは比較的大きいと思われることなどから、一般に防衛共生体は進化的に不安定な地位にあり、宿主個体群中で定常的に維持されず、ゲノムが大幅に縮小することもない(>>100万塩基対)と考えられてきました。今回私たちは、この常識を覆す、進化的にきわめて安定で宿主と一体化した特徴を示す「防衛共生体」を、ミカンキジラミから発見しましたので、これについてご紹介します。

図2: (左上) カルソネラのゲノム。(右) プロフテラの染色体。二次代謝関連遺伝子群(dipA-dipT)が、ゲノム上の広い領域を占めることが分かる。(左下) プロフテラのプラスミド。
(クリックで拡大します)
ミカンキジラミの複合共生系
ミカンキジラミ (Diaphorina citri)(図1)は、カンキツグリーニング病を媒介することで世界のカンキツ産業に大きな被害を与えている、重要な農業害虫です。腹部の体腔内に「バクテリオーム」とよばれる共生専用の器官を持ち、その表層の細胞内に、キジラミ類に普遍的に存在する「Candidatus Carsonella ruddii (ガンマプロテオバクテリア綱、以下カルソネラ_DC)」、バクテリオームの内腔内に、これまで同種でのみ見つかっている「Candidatus Profftella armatura (ベータプロテオバクテリア綱、以下プロフテラ)」を収納しています。私たちの先行研究により、カルソネラ以外の共生細菌をもたないキジラミPachypsylla venustaのカルソネラ(カルソネラ_PV)のゲノムは、葉緑体ゲノムと同レベルの16万塩基対にまで縮小し、細胞生物として際立って乏しい遺伝子組成を持つことが明らかとなり(Nakabachi et al, 2006)、カルソネラのゲノム進化や、プロフテラが加わることにより新たに付与される共生の機能に興味が持たれました。これらの解明をめざし、まずミカンキジラミ共生細菌2種の全ゲノム塩基配列を決定しました(図2)。
カルソネラはゲノム縮小の極端に進んだ栄養共生体
その結果、カルソネラ_DCのゲノム(174,014塩基対)は、カルソネラ_PV(159,662塩基対)と同様きわめて小さく、ゲノム全体の構造がカルソネラ_PVとの間で良く保存されており、その遺伝子組成は、キジラミのえさである植物師管液に乏しい必須アミノ酸の合成に特化したものであることが分かりました。この事実は、カルソネラ_DCがミカンキジラミに不足する栄養を補う「栄養共生体」であることを示します。先に述べたように、栄養共生体の多くは宿主の生存に不可欠であり、垂直感染のみにより永続的に受継がれ、その過程でゲノムサイズが縮小する傾向を示します。カルソネラはそのゲノム縮小の最も極端な例のひとつと言えます。
前代未聞! 極小ゲノムを持つ防衛共生体、プロフテラ
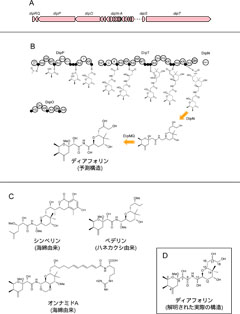
図3: ディアフォリンの構造解析。(A) プロフテラゲノム上のディアフォリン合成系遺伝子群。(B) 遺伝子組成と遺伝子構造に基づく代謝経路・産物構造の推定。(C) ディアフォリンの類縁体。(D) NMR・質量分析などにより解明されたディアフォリンの実際の構造。
(クリックで拡大します)
一方、プロフテラのゲノムサイズは464,857塩基対で、カルソネラには及ばないものの、やはりきわめて小さいものでした。これは、プロフテラが進化的時間にわたり、宿主キジラミとの間で、互いになくてはならない緊密な相利共生関係にあることを示唆します。これまで知られていた、同様のゲノムサイズを持つ共生細菌は、いずれも宿主に栄養分を供給するための遺伝子群を備えた「栄養共生体」でした。ところが、プロフテラのゲノムを調べると、こうした遺伝子はごくわずかで、その代わりに、毒などの二次代謝産物の合成に関わるらしい遺伝子群がゲノム全体の15%もの広い領域を占めました(図2)。その遺伝子レパートリー(図3A)やそれぞれの遺伝子の構造を手がかりとして、産物の構造を推定し(図3B)、この情報にもとづいて、ミカンキジラミから標的化合物を単離、定量したところ、予測された物質が、実際にミカンキジラミの体内に高濃度で含まれることが明らかとなりました。さらに、さまざまな手法を適用することで、化合物の詳細な構造を決定したところ、プロフテラは、ハネカクシのもつ強力な細胞毒「ペデリン」(図3C)と良く似た構造の新規化合物(図3D)を合成することが明らかとなり、「ディアフォリン」と名付けました。また、ディアフォリンを2種類の培養細胞に与えたところ、これらの細胞を死滅させる強い毒性が示されました(図4)。さらに、ディアフォリン合成系の遺伝子群について分子系統解析を行うと、これらが「水平転移」により獲得されたものであり、その多くが、ペデリン合成関連遺伝子群と単系統群を形成することが分かりました。また、世界の9つの個体群の合計806頭(雄: 409頭、雌: 397頭)について、共生細菌およびディアフォリン合成系遺伝子の存否を調べたところ、806頭すべての個体がカルソネラとプロフテラの両者に感染していること、およびすべての個体中にディアフォリン合成系遺伝子が存在することが確認されました。
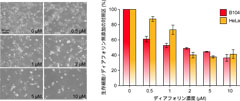
図4: (左) 各濃度のディアフォリン添加48時間後のラットB104細胞。(右) 各濃度のディアフォリン添加48時間後のラットB104細胞とヒトHeLa細胞の、対照区に対する生細胞数の比。
(クリックで拡大します)
以上のことからプロフテラは、あらゆる個体に存在し、宿主の共生器官に収納されて垂直感染により受継がれ、ゲノムサイズはわずか 46万塩基対と多くの栄養共生体のゲノムさえ下回り、宿主動物と一体化したオルガネラのような特徴を示しながら、毒を合成することで宿主を天敵から守る「防衛共生体」 であると考えられ、これまでの防衛共生についての常識を覆しました。
今後の展開
現在私たちはミカンキジラミに近縁なキジラミの共生細菌叢の解析を進めており、プロフテラの姉妹種が見つかり始めています。今後はこうした細菌の解析を進めることで、プロフテラの特異な進化過程を明らかにして行きたいと考えています。
引用文献
- Nakabachi A, Ueoka R, Oshima K, Teta R, Mangoni A, Gurgui M, Oldham NJ, van Echten-Deckert G, Okamura K, Yamamoto K, Inoue H, Ohkuma M, Hongoh Y, Miyagishima SY, Hattori M, Piel J, Fukatsu T. (2013) Defensive bacteriome symbiont with a drastically reduced genome. Curr. Biol. 23(15): 1478-1484.
- Nakabachi A, Yamashita A, Toh H, Ishikawa H, Dunbar HE, Moran NA, Hattori M. (2006) The 160-kilobase genome of the bacterial endosymbiont Carsonella. Science 314(5797): 267.
- 該当記事のプレスリリース(豊橋技術科学大学)
- 該当記事のNature Research Highlightsによる紹介 “Citrus pest's little helper”
応用動物学/昆虫学最新トピック
プロの研究者でもまだ知らないような、出来たてホヤホヤの最新研究成果を分かりやすくお伝えします。
日本応用動物昆虫学会(応動昆)
「むしむしコラム・おーどーこん」は、日本応用動物昆虫学会電子広報委員会が管理・運営しています。