2008年09月12日掲載 【トレハロース: 干からびたネムリユスリカ幼虫が蘇生する鍵分子】
乾燥に強い多くの昆虫はクチクラを厚くするなど身体から水分を失うことを回避していますが、ネムリユスリカは昆虫で唯一全く異なった戦略で乾燥ストレスに対応しています。すなわち身体が完全に干からびても死なない機構を獲得しているのです。
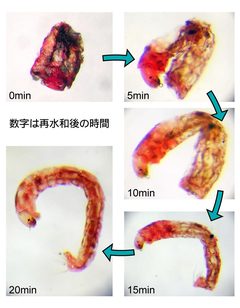
図1: 吸水により発育再開するネムリユスリカ幼虫
(クリックで拡大します)
ネムリユスリカはアフリカの半乾燥地帯の岩盤の窪みにできた小さな水たまりに生息しており、乾季になると水たまりは干上がり幼虫もカラカラに乾いてしまいますが、雨季がきて雨が降ると吸水し1時間ほどで発育を再開します(図1)。ネムリユスリカ幼虫の持つ細胞や器官、すなわち脳を含む神経内分泌器官、消化器官、脂肪体とマルピーギ氏管、血球、骨格筋などの組織や細胞はほぼ完全に脱水した無代謝の状態(クリプトビオシス)で、しかも常温で長期間(最長記録は17年)保存されていることになります。このネムリユスリカの驚異的な乾燥耐性の仕組みを明らかにし、細胞や臓器の常温保存技術に応用できればと思っています。
トレハロースの生体成分保護機能
我々人間の細胞は、細胞レベルでは、意外にも乾燥に強く50%の脱水にも耐えます。しかしそれ以上の脱水が進むと活性酸素の発生によるタンパク質や脂質の酸化、DNA切断など生体成分の損傷がアポトーシスを誘導し細胞死に至ります。一方、ネムリユスリカを含む多くのクリプトビオシス生物は、水の代わりに生体成分を保護する物質(適合溶質)としてトレハロースを蓄積したり、活性酸素の発生を抑制する抗酸化酵素を合成することで、ストレスによる損傷を回避しています。トレハロースについては乾燥重量当たり20%に相当する量を蓄積し生体成分を保護していますが、物理化学的な保護機能については、2つの仮説が提唱されてきました。ひとつは水置換説で、完全脱水状態においてトレハロースは細胞膜やタンパク質の表面に直接水素結合し、結果的に結合水の代理をするという考え方。もうひとつは、ガラス状態説で、トレハロースの水溶液が、脱水に伴い、流動性を失いガラス化し、細胞膜やタンパク質は、一種のマイクロカプセルの中に閉じ込められる形になり、それらの高次構造はそのまま保護されるという仮説です。最近これらの仮説がどちらも妥当であることを東京工業大学の櫻井先生のグループと共同でネムリユスリカを用いて物理化学的に証明することができました(Sakurai et al. 2008)ので紹介します。
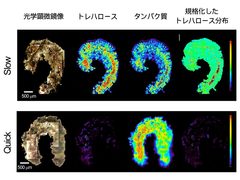
図2: 顕微FTIRによるトレハロースの体内局在解析
(クリックで拡大します)
トレハロースの幼虫体内局在
クリプトビオシスする生物はワムシやクマムシのように概して微小であること、トレハロースは非還元糖ゆえに染色が困難なことなどの理由でクリプトビオシス状態の生物の体内トレハロース局在は不明でした。我々はネムリユスリカ幼虫の優位性(クリプトビオシス生物の中で最大)を利用し、トレハロースの幼虫体内局在を明らかにしました。ネムリユスリカ幼虫を48時間かけてゆっくり乾燥させた幼虫(これをSlowと呼ぶことにします)は大量のトレハロースを合成しますが、幼虫体内のどこに局在しているのかは不明でした。顕微FTIR(顕微鏡と赤外線分光を合体させた装置)により、それが体内全体に均一に分布していることが赤外吸収スペクトル測定によって判明しました(図2)。一方、数時間で急速に乾燥した幼虫(これをQuickと呼びます)は、わずかなトレハロースしか合成できず、水に戻しても蘇生しませんでした。
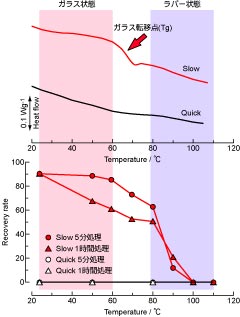
図3: DSC解析によるガラス転移温度(上)と熱耐性(下)との関係
(クリックで拡大します)
トレハロースのガラス化
ガラスとは、結晶とは異なり分子や原子が不規則な空間配置を維持したまま固形になった状態を指します。日常目にする窓ガラスはシリコンなど無機物質が"ガラス化"した状態です。バーナー等でガラスを加熱すると、やがて分子や原子の動きが急激に活発になり、ガラスは軟化します。これをガラス転移と言い、その境となる温度をガラス転移温度(Tg)と言います。近年、トレハロースなどの糖類も温度や湿度等の条件に応じてガラス化することが広く知られるようになりました。ガラスには分子などの配列に規則性は要求されないため、細胞膜やタンパク質が共存しても互いに分離することなくトレハロースは固化します。その結果としてガラス状態のトレハロースが生体物質をカプセル状に包み込み、乾燥ストレスから保護していると予想されました。そこで蘇生率の異なる両サンプルを示差走査熱量計(differential scanning calorimeter, DSC)を用いて0~100℃の範囲で5℃/min.でスキャンし、ガラス転移温度を測定したところ、蘇生可能なトレハロースを蓄積しているSlowでは明瞭なガラス転移挙動が観測されましたが(図3上 矢印)、トレハロースをほとんど持たない蘇生不可能なQuickではガラス転移が確認されませんでした。Slow幼虫に対して80℃以上の温度処理をすると、再水和後の蘇生率が急速に低下しました(図3下)。トレハロースの生体成分の保護機能は、ガラス状態からラバー状態に相転移して失われたことになります。ちなみに、転移の開始点は56℃、転移中点は65℃、終了点は72℃でした。自然界では、ネムリユスリカが棲息する岩盤の表面温度は日中60℃にも達しますが、乾燥幼虫のガラス転移温度の終了点はそれ以上であることから、クリプトビオシスは問題なく維持されます。トレハロースは糖類の中でもガラス化しやすく、ガラス転移温度が高いという性質を持ちます。このトレハロースを適合溶質として選択したことが、ネムリユスリカが乾期に生き残ることに成功した理由の一つかもしれません。
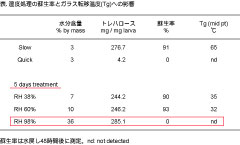
表: 湿度処理の蘇生率とガラス転移温度(Tg)への影響
(クリックで拡大します)
湿気はガラス状態を保持するには不利な条件であることが物質科学の分野でよく知られています。そこでSlowサンプルがガラス状態であることと、それが蘇生可能であることの関係をより明確にするため吸湿試験も行いました(表)。例えば、ネムリユスリカ乾燥幼虫(相対湿度5%に保存)を相対湿度98%の条件に移し5日間置くと、含水量3%だったのが吸水して36%になり、DSCで計測するとガラス転移曲線が確認されませんでした。体内に大量のトレハロースを蓄積していても、それがガラスではなくなると生体成分の保護機能は失われ、幼虫は水に戻しても蘇生しませんでした。ネムリユスリカのクリプトビオシスはアフリカの半乾燥地帯の乾季でのみ成立する現象かもしれません。
トレハロースの水置換
代謝活動が行われている通常の生体では、細胞膜は潤沢な水に覆われ、流動性に富んだ柔軟性をもった液晶状態にあります。乾燥により細胞膜表面の水分(結合水)が失われると、細胞膜は柔軟性を失い、ゲルと呼ばれる固化した状態に変化します。クリプトビオシス幼虫の細胞膜が液晶状態を維持していることが確認できれば、水置換仮説の証明をしたことになります。分光分析の結果、Slowの細胞膜は、乾燥した状態にあるにも関わらず通常の生命活動状態のときと同様に、常温でも高い流動性をもった液晶状態に保たれていることが分かりました。一方、Quickでは、常温での細胞膜の流動性は一部失われ、部分的にゲルと呼ばれる固い状態になっていました。ここで細胞膜を構成するリン脂質を爪楊枝に例えて補足説明をします。円筒形の容器に100本ほどの爪楊枝がぎっしり入った状態を想像してください。そこから20本ほど爪楊枝を抜くと、上から指を1本差し込めるくらい爪楊枝は横への移動ができる状態になります。これが液晶状態で、水和した細胞の細胞膜はこのような状態にあります。Slowの細胞膜というのは、本来水が存在しているところにトレハロースが置換しガラス化しながら液晶状態を保持しているものと考えられます。一方、爪楊枝がぎっしり詰まって縦にも横にも動かない状態がゲルです。Quickでは、乾燥と再水和の過程で、液晶からゲルへと相変異が生じます。その際に、細胞膜上の複数箇所でゲル化が生じると液晶とゲルの異なる相の境界線は亀裂ができますのでこれが細胞膜の損傷の原因となります。トレハロースの水置換は幼虫の脱水の過程の途中で、ガラス化は終盤で起こります。
おわりに
乾燥ストレスは脱水に伴う塩ストレスなどと共に活性酸素を発生させ、それがタンパク質や脂質の酸化やDNA切断をもたらし細胞死(アポトーシス)や細胞のガン化を引き起こします。ネムリユスリカの幼虫体内では、乾燥や塩ストレスの刺激によって生体成分を保護する機構が発動します。しかし生体成分の損傷は完全に回避できないことが最近の研究でわかってきました。どうやらネムリユスリカはその損傷を蘇生時に修復しているようです。クリプトビオシス現象の解明研究は新たな方向に展開し始め、ますます面白くなってきました。
参考文献
- Sakurai M., Furuki T., Akao K., Tanaka D., Nakahara Y., Watanabe M., Kikawada T., Okuda T. (2008) Vitrification is essential for anhydrobiosis in an African chironomid, Polypedilum vanderplanki. Proc. Nat. Acad. Sci. USA 105(13): 5093-5098
« トレハロース: 干からびたネムリユスリカ幼虫が蘇生する鍵分子 | トピック一覧 | トレハロース: 干からびたネムリユスリカ幼虫が蘇生する鍵分子 »
応用動物学/昆虫学最新トピック
プロの研究者でもまだ知らないような、出来たてホヤホヤの最新研究成果を分かりやすくお伝えします。
日本応用動物昆虫学会(応動昆)
「むしむしコラム・おーどーこん」は、日本応用動物昆虫学会電子広報委員会が管理・運営しています。